




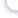
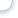


Аквакультура В.И. КОЗЛОВ, А.Л. НИКИФОРОВ-НИКИШИН, А.Л. БОРОДИН. Потребность форели в кислороде
Специфическое потребление кислорода радужной форелью
при кормлении гранулированным комбикормом, мг/кг х ч (О2сп)
| Масса рыбы, г | Температура воды, 0С | |||||||||
| 0,08 | - | - | - | - | - | |||||
| 0,2 | - | - | - | - | - | - | ||||
| 0,5 | ||||||||||
| 1,0 | ||||||||||
| 5,0 | ||||||||||
| Масса рыбы, г | Температура воды, 0С | |||||||||
| 0,08 | - | - | - | - | - | |||||
| 0,2 | - | - | - | - | - | |||||
| 0,5 | ||||||||||
| 1,0 | ||||||||||
| Масса рыбы, г | Температура воды, 0С | |||||||||
| 0.08 | - | - | - | - | - | - | ||||
| 0.2 | - | - | - | - | - | - | ||||
| 0,5 | ||||||||||
| 1,0 | ||||||||||
При выращивании радужной форели, как одного из основных объектов индустриального рыбоводства при температуре воды 14 –18 0С принято, что 90% кислорода используется для дыхания, а 10% - для окисления органических веществ, находящихся в рыбоводной емкости (остатки корма, экскременты, органические взвеси в поступающей воде и др.).
Учитывая данные о поступлении и расходе кислорода, может быть составлено следующее уравнение баланса кислорода в рыбоводной емкости (для радужной форели):
(1) 0,9 (О2'' - О2') n х V= О2сп х Р, где
О2'' - О2'– содержание растворенного кислорода на втоке и вытоке, мг/л;
n – смена воды в бассейне, раз в час;
V – рабочий объем рыбоводной емкости, м3;
О2сп – специфическое потребление кислорода радужной форелью, мг/кг х ч;
Р – общая масса рыбы в рыбоводной емкости, кг
Левая часть уравнения кислородного баланса показывает количество растворенного кислорода в рыбоводной емкости при определенной температуре воды, которое может быть использовано рыбой для дыхания. Коэффициент 0,9 показывает, что 90 % кислорода идет на дыхание, а 10 % - на окисление органических веществ в бассейне. Величина О2’ на вытоке не должна опускаться ниже 7 мг/л для форели, поскольку ниже этой величины у форели наступает ухудшение обмена. Для других рыб, например для карпа, минимальная величина О2’ на вытоке может составлять 5 мг/л. Первая часть уравнения показывает специфическое потребление кислорода всей рыбой при определенной температуре воды и определенной индивидуальной массе рыбы в условиях кормления сухим гранулированным кормом по кормовым таблицам.
Под плотностью посадки понимается количество рыбы на единице площади и объема воды, которую можно выразить формулой:
(2) СР=P/V, где
СР – плотность посадки рыбы, кг/м3; Р – общая масса рыбы, кг;
V – объем рыбоводной емкости, м3 (рабочий объем).
Пользуясь уравнением (1) и формулой (2) и выражая рабочий объем в литрах, можно рассчитать плотность посадки рыбы при заданной проточности:
( 3 ) СР= 0,9 (О2'' – О2 ') 1000 · n/ О2 сп.,где
n - заданная величена смены воды в бассейне, раз в час (интенсивность обмена).
Интенсивность водообмена n непосредственно связанна с расходом воды:
(4) N = n V / 3600 , где
N - расход воды, л/с
V - объем рыбоводной емкости, м3.
Следовательно, общий расход воды, необходимый для выращивания определенного количества рыбы с определенной индивидуальной массой при конкретной температуре составит:
(5) N = РО2сп (О2'' - О2') 0,9
1. Например: При температуре воды 5 0С содержание растворенного кислорода при 100 %- ном насыщении составляет 12,81 мг/л, а при 15 0С – 446 мг/кг х ч. Рассчитываем по формуле 3 .
Максимальная плотность посадки при 5 и 15 0С составит:
СР5 = 0,9 (12,81 – 7,0 ) 1000 х 3 / 155 = 101,2кг/м3
СР15 = 0,9 (10,18 – 7,00) 1000 х 3 / 446 = 19,2кг/м3
При этом в обоих случаях расход воды составит:
N = 1000 х 3 / 3600 = 0,83 л/с
Таким образом, повышение температуры воды при сохранении той же проточности вызывает снижение плотности посадки рыбы. Для увеличения плотности посадки необходимо повысить водообмен.
2. Например: Определить максимальную плотность посадки радужной форели массой 100 г при температуре воды 5 и 15 0С, интенсивности водообмена 3 раза в час, содержании кислорода на вытоке 7 мг/л. Расчеты следует проводить по формуле 3.
При температуре 5 0С содержание растворенного кислорода при 100 %-ном насыщении составляет12,81, а при температуре 15 0С – 10,18 мг/л. Специфическое потребление кислорода форелью при 5 0С – 126 мг/кг х ч, а при 15 0С – 360 мг/кг х ч. Таким образом, максимальная плотность посадки составит:
СР5 = 0,9 (12,81 – 7,0) х 1000 х 3 / 126 = 124,5 кг/м3
СР15 = 0,9 (10,18 – 7,0 ) х 1000 х 3 / 360 = 23,8 кг/м3
При этом в обоих случаях расход воды составит 0,83л/с:
N = 1000 х 3 / 3600 = 0,83 л/с
Плотность посадки рыбы при одном и том же водообмене и уровне кислорода тем выше, чем больше индивидуальная масса рыбы и ниже температура воды.
При выращивании рыбы на предприятиях индустриального типа следует создавать оптимальный режим температуры и насыщение воды кислородом. Это достигается использованием нагретой технологической воды тепловых электростанций или применением специальных установок для нагрева. Уровень кислорода в рыбоводных емкостях должен быть равен 100 % - ному насыщению (см. табл. 1) или близким к нему. Природная вода после подогрева не содержит такое количество кислорода, поэтому следует применять методы аэрации воздухом или чистым кислородом, причем последнее предпочтительнее из-за более высокой эффективности.
Увеличение интенсивности водообмена с целью улучшения газового состава имеет ограничения, объясняемые физическим воздействием течения на рыб и значительным расходом энергии на удержание тела в потоке.
Среди методов определения плотности посадки культивируемых рыб в условиях индустриального рыбоводства привлекает внимание метод, основанный на том, что концентрация рыбы или плотность посадки в единице рыбоводной емкости определяется количеством кислорода, необходимого для окисления суточной нормы корма.Как известно, спокойная, не питающаяся рыба потребляет меньше кислорода, чем активная, питающаяся. Потребление кислорода резко возрастает у питающейся рыбы за счет усиления обмена, окисления съеденного корма и выделения продуктов обмена. Возможное количество корма, которое может быть использовано рыбой при конкретном количестве кислорода может быть вычислено следующем образом (Haskell, 1955):
(6) Х = (О2'' - О2') 1,44 · N / 220, где
Х - количество корма, кг/сут.; О2''- начальное содержание кислорода в притекающей воде, мг/л; О2' - конечное минимальное содержание кислорода в вытекающей воде, мг/л; N - количество воды, подаваемой в данную рыбоводную емкость, л/мин; 1,44 - количество воды в сутки при интенсивности подачи 1л/мин, т; 220 - необходимое количество кислорода для усвоения рыбой 1 кг гранулированного корма с калорийностью 2600-2800 ккал/г (вычислено на основаниях эмпирических данных за 10 лет работы питомника Мак Нении, США).
Например: Содержание кислорода в притекающей воде равно 10 мг/л, количество подаваемой воды - 150 л/мин. Рассчитать количество корма для форели в сутки. Расчеты следует проводить по формуле 6.
Х= (10 – 7) 1,44 · 150/ 220 = 2,94 кг,
то есть при данных условиях в выростную емкость можно вносить 2,94 кг корма в сутки.
Определив количество корма, которое может быть использовано при данном количестве кислорода, рассчитываем возможное количество рыбы в рыбоводной емкости и плотность посадки. При этом используют кормовые таблицы, например, таблицы ВНИИПРХ (Канидьев, Гамыгин, 1983), в которых показана суточная норма кормления форели в зависимости от массы тела и температуры воды, то есть,
(7) Р = Х./ Нсут , где
Р - возможная общая масса рыбы в рыбоводной емкости, кг;
Х - количество корма в сутки, кг;
Нсут- суточная норма корма в % массы рыбы.
Например: Температура воды, подаваемой в бассейн рыбоводного предприятия индустриального типа, равна 10 0С, масса рыбы 12 г, следовательно (по кормовым таблицам), для сухих гранулированных кормов суточная норма составит 2,6 % к массе рыбы. Расчеты следует проводить по формуле 7.
Р = 2,94 /0,026 = 113,1 кг или 113,1 кг / 12 г = 9423 шт
Как видно, метод расчета плотности посадки рыбы, предложенный Х. Уиллаби (1968), основан на потребности в кислороде в зависимости от количества вносимого корма. Эта потребность в кислороде определена эмпирически и фактически учитывает зависимость потребления кислорода от температуры воды, размера рыбы и качества корма. Метод учитывает также и влияние продуктов обмена на способность рыбы использовать кислород в данных условиях кормления.
Таким образом, этот метод достаточно универсален. Однако он требует подробных данных о величине суточного рациона в зависимости от температуры воды и массы рыбы. К настоящему времени эта зависимость изучена весьма тщательно, в основном для лососевых и карповых рыб (Канидьев, Гамыгин, 1986, Гамыгин и др., 1988). Она учитывает изменение физиологической активности при разной температуре, следовательно, учитывает изменение общего обмена. Так, при температуре 5 0С суточный рацион радужной форели массой 2-5 г составляет 2,2 %, при температуре 10 0С – 3,3 %, а при 15 0С – 4,9 % от массы рыбы (Канидьев, Гамыгин, 1977).
Суточный рацион имеет обратную связь с массой тела рыбы. Если суточная норма для молоди лососевых массой 2 г при температуре 10 0С равна 4,2 %, то для молоди массой 12-25 г – вдвое меньше.
**Температура воды на рыбоводных предприятиях индустриального типа проявляется во взаимодействии с определенным газовым составом воды. Вода поверхностных источников всегда в той или иной мере насыщена кислородом, уровень которого в первую очередь зависит от температуры (табл.А). Существует четко выраженная зависимость между уровнем растворенного в воде кислорода и белковым, жировым и углеводородным обменом у рыб. Среди культивируемых рыб лососевые являются наиболее оксифильными, карповые – наименее оксифильными. Пороговая концентрация кислорода с возрастом понижается. Так, свободные эмбрионы стальноголового лосося и радужной форели погибают при содержании кислорода 2,2-2,7 мг/л, годовики – 2,0-2,4 мг/л, а двухлетки – 1,5-2,0 мг/л, тогда как соответствующие возрастные группы карпа погибают лишь при содержании кислорода примерно вдвое более низком. Осетровые рыбы занимают среднее положение.
Таблица А. Нормальное насыщение пресной воды кислородом при нормальном
poisk-ru.ru
КИСЛОРОД И ЕГО ПОТРЕБЛЕНИЕ РЫБОЙ
Просмотров: 5421
Для всех культивируемых водных объектов необходим кислород, который им приходитсяизвлекать из воды. Основным источником пополнения кислорода в природной воде являетсяатмосфера и фотосинтезирующие растения. В атмосфере содержится 21% кислорода, а содер-жание кислорода в воде зависит от различных факторов, в частности, от температуры воды исодержания в ней ионов хлора. Для каждой температуры и солености (хлорности) может бытьустановлено равновесное 100% насыщение атмосферным кислородом. В холодной пресной во-де насыщение выше, чем в более теплой и насыщенной ионами хлора. Равновесное насыщениевесьма важный показатель, по которому оценивается несущая способность воды по кислороду.Для практического пользования эта информация представлена в виде табл.12.Таблица 12.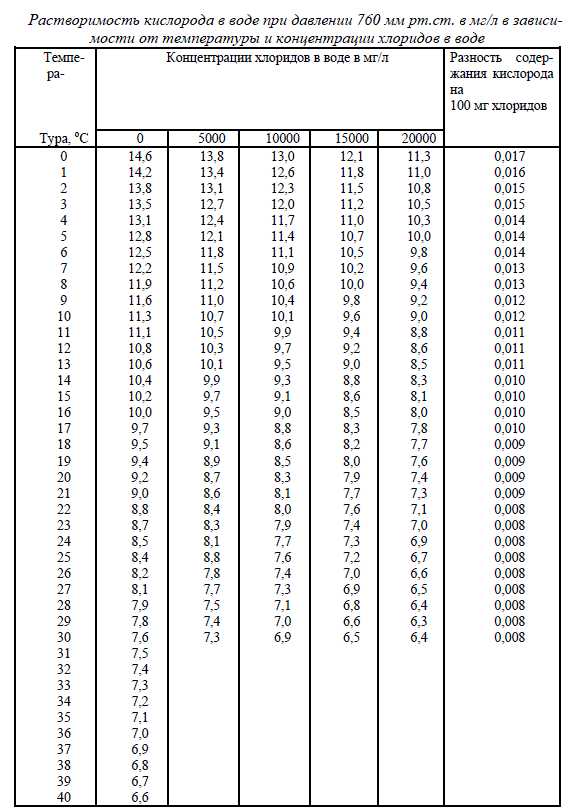
Если по каким-либо причинам насыщение воды в водоеме оказывается ниже равновесногонасыщения, то кислород воздуха переносится в воду из-за возникающего градиента концентра-ций.
Скорость процесса переноса кислорода тем выше, чем выше градиент концентрации, тоесть чем выше разница между равновесным и фактическим насыщением воды. При приближе-нии фактического насыщения воды к равновесному скорость процесса падает. При пресыщениеводы кислородом выше равновесного насыщения идет процесс потери кислорода из воды(диффузия). Процессы насыщения и диффузия идут через поверхность раздела сред во-да/воздух, поэтому, чем больше поверхность раздела, тем интенсивнее идет процесс. Поверх-ность раздела может быть искусственно увеличена за счет разбрызгивания или барботированиявоздухом. Создание турбулентного течения воды также увеличивает скорость процесса, так какувеличивается градиент концентрации на границе раздела при перемешивании слоев воды оразличной степенью насыщения. Процессы диффузии кислорода в неподвижной воде настолькомало влияют на уровень насыщения различных слоев воды, что ими можно пренебречь.Фотосинтез - второй основной источник кислорода для водных систем. Кислород являетсяпобочным продуктом фотосинтеза растений. Однако, фотосинтез и продуцирование кислородапроисходят только при наличии света. Те водоросли, которые в дневное время выделяют ки-слород, в ночное время его поглощают. В солнечные дни количество кислорода, выделяемоговодорослями в воду, может быть настолько большим, что уровень насыщения воды значитель-но превышает уровень равновесного насыщения. В этом случае кислород выделяется из воды ватмосферу. Известны случаи пресыщения воды до 300% от равновесного значения. Макси-мальная концентрация кислорода бывает в середине дня или несколько позднее, а минимальнаяконцентрация - на рассвете.Потребление водорослями кислорода в темное время суток может снизить его концентрациюв воде до критического для культивируемых объектов значения. Особую опасность представ-ляют тихие безветренные ночи, когда пополнение воды кислородом воздуха затруднено. Кон-троль концентрации кислорода в открытых водоемах следует проводить в утренние часы.НОРМА КОНЦЕНТРАЦИИ КИСЛОРОДА В ВОДЕ РЫБОВОДНЫХ УСТАНОВОК.В естественных открытых водоемах, в результате процесса фотосинтеза водорослей, верхняяграница насыщения может оказаться равной 300% от равновесного насыщения. При этом неотмечено гибели рыбы или каких-либо других отрицательных последствий.Снижение концентрации кислорода до 60% от равновесного насыщения практически не ска-зывается на росте и использовании кормов форелью и карпом. Временное снижение концентра-ции кислорода ниже 60% приводит к стрессам на несколько дней, рыба прекращает брать корм.При дальнейшем снижении концентрации кислорода наступает гибель рыбы. Нижние порого-вые концентрации кислорода зависят от вида и массы тела рыбы, а также от температуры воды.Пороговые концентрации кислорода являются важной информацией при выполнении операцийпо перевозке рыбы. В этот момент рыба не питается, а единственной задачей рыбовода являетсясохранение рыбы в живом виде. Пороговые концентрации кислорода в воде для различных ви-дов рыб приведены в табл.13.Оптимальные концентрации кислорода для рыбы подбираются в зависимости от стадии ееразвития; икра, личинки, молодь, товарная рыба. На стадии инкубации икры требования куровню концентрации кислорода жестче, чем на других стадиях. Например, при инкубации ик-ры карпа снижение концентрации кислорода до 6,5 мг/л, что составляет 70% равновесного на-сыщения, оказывается летальным. В период усиленного развития икринки, во время закладкиосновных органов и тканей зародыша, даже 100% насыщение воды 8,5 мг/л становится крити-ческим, часть икры погибает. Повышение концентрации кислорода до 25 - 35 мг/л не сказыва-ется на скорости развития икры. Однако, при столь высокой концентрации кислорода уменьша-ется отход икры. В дальнейшем развитии молодь, полученная из икринок, инкубированных приповышенной концентрации кислорода, развивается и набирает массу быстрее молоди, по лу-ченной из икры, инкубированной при 100% насыщении воды.45Таблица 13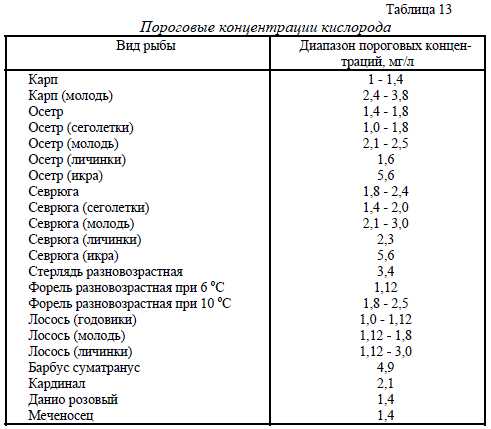
На стадии личинки, когда организм рыбы претерпевает морфологические преобразования,требования к уровню концентрации кислорода не менее жесткие. Концентрация кислородадолжна поддерживаться на уровне 100% насыщения, хотя более высокие концентрации не при-носят очевидного вреда, если они не связаны с газопузырьковой опасностью.На более поздних стадиях развития молоди и выращивания товарной рыбы оптимальныеконцентрации кислорода лежат в пределах от 90 до 200% насыщения. Дальнейшее увеличениеконцентрации эффекта не дает. Очевидно, что при высоких концентрациях кислорода усвоениекорма рыбой идет эффективнее, а затраты энергии на дыхание и поиск зон с повышенной кон-центрацией кислорода сводятся к минимуму. О недостатке кислорода можно судить по поведе-нию рыбы. На недостаток кислорода указывает концентрация рыбы на вводе воды в бассейн,выпрыгивание, скопление рыбы в верхних слоях воды.ПОТРЕБЛЕНИЕ КИСЛОРОДА РЫБОЙ.Потребление кислорода рыбой зависит от стадии развития, массы тела, температуры и коли-чества потребленного корма. По мере развития эмбриона потребность в кислороде возрастает.При инкубировании икры в условиях цеха потребность в кислороде удовлетворяется за счет во-ды, протекающей через инкубатор. При перевозке икры на стадии глазка потребность в кисло-роде покрывается тем запасем кислорода, который помещен в контейнер для перевозки икры. Сростом температуры потребление кислорода эмбрионами растет. Рост потребления кислородаэмбрионами осетровых приведен в табл.14.Таблица 14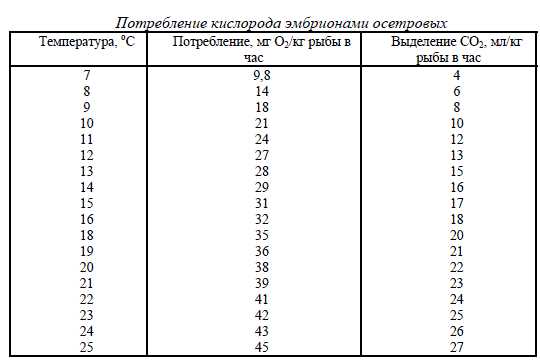
При переходе личинок на питание внешним кормом по мере развития и роста рыбы удельноепотребление кислорода на единицу массы рыбы падает. Одновременно сохраняется зависи-мость уровня удельного потребления кислорода в функции температур в пределах температур,возможных для разведения. Эта зависимость иллюстрируется потреблением кислорода лосося-ми и форелью в диапазоне температур от 7,2 до 21,1 оС для рыб массой до 1000 г. (рис.21). Рис. 21. Потребление кислорода в зависимости от массы рыбы и температуры.Потребление кислорода в течение суток неравномерно. Максимальное потребление ки-слорода у форели наблюдается чрез 10 - 15 мин после внесения корма и снижается через 1 - 1,5часа. Уровень потребления кислорода колеблется в пределах 1,2 - 2 раза. Аналогичная картинанаблюдается при кормление карпа. По этой причине все данные о потреблении кислорода при-водятся в расчете на сутки. Для практических расчетов потребности кислорода для рыбы расчетведется по потребленному ей корму. В пределах температур оптимальных для форели потреб-ление кислорода пропорционально потреблению корма. На 1 кг сухого гранулированного кормарыбой расходуется 0,22 кг кислорода. Такой расчет применим только для условий нормальноготехнологического процесса выращивания рыбы. Действительно, при прекращение кормлениярыбы потребление кислорода не станет равным нулю.
Рис. 21. Потребление кислорода в зависимости от массы рыбы и температуры.Потребление кислорода в течение суток неравномерно. Максимальное потребление ки-слорода у форели наблюдается чрез 10 - 15 мин после внесения корма и снижается через 1 - 1,5часа. Уровень потребления кислорода колеблется в пределах 1,2 - 2 раза. Аналогичная картинанаблюдается при кормление карпа. По этой причине все данные о потреблении кислорода при-водятся в расчете на сутки. Для практических расчетов потребности кислорода для рыбы расчетведется по потребленному ей корму. В пределах температур оптимальных для форели потреб-ление кислорода пропорционально потреблению корма. На 1 кг сухого гранулированного кормарыбой расходуется 0,22 кг кислорода. Такой расчет применим только для условий нормальноготехнологического процесса выращивания рыбы. Действительно, при прекращение кормлениярыбы потребление кислорода не станет равным нулю.
biblio.arktikfish.com
При этом на 1 л/с можно довести плотность посадки до 27,3 кг/м3 (табл.11). Таблица 11 Максимальная плотность посадки радужной форели при расходе воды 1 л/с и температуре воды 110С
Товарная форель массой 200 г в зависимости от температуры воды потребляет различное количество растворенного кислорода и требует разного расхода воды (табл. 12). Таблица 12 Потребность радужной форели массой 200 г в кислороде и воде в зависимости от температуры и содержания растворенного кислорода
Сотрудниками ВНИИПРХ на основе анализа прогрессивных технологий и собственных выводов разработаны нормативы, соответствующие интенсивным методам разведения и выращивания форели. В них рекомендуется смена воды в выростных и нагульных прудах за 20-30 мин, а вмальковых басссейнах за 8-10 мин, что может обеспечить получение 50-60 кг/м3 рыбопродукции. Максимальная плотность посадки молоди форели существенно изменяется от средней массы, температуры воды и кратнсти водообмен (табл. 13). Таблица 13 Максимальная плотность посадки молоди форели при разной температуре воды и различном водообмене, кг/м3
По нормативам Гидрорыбпроекта (Каспин и др., 1976) на 1 кг форели расход воды планировался в 3-4 раза меньше, чем по нормам ВНИИПРХа, соответсвенно были занижены показатели плотности посадки и общий уровень рыбопродукции. В самом крупном форелевом хозяйстве (США) с площади прудов в 4 га получают 600 т форели или при общем расходе воды 4 м3/с по 150 т/га (Бардач и др., 1978). При высоких плотностях посадки здесь получают 132 кг форели на 1л/с в год. Найдено, что при увеличении проточности в бассейнах и прудах уровень интенсификации форелеводства может быть повышен по сравнению с существующим (10 кг/м3) в 16 раз без угнетения роста рыбы и увеличения затрат корма на прирост. Отмечено также, что уровень интенсификации в форелеводстве наряду с другими факторами, особенно кормлением, во многом определяется уровнем водоснабжения или водообмена в рыбоводных емкостях и качеством поступающей в них воды, прежде всего по содержанию кислорода. В современных форелевых прудовых хозяйствах форель выращивают при относительно низком уровне водообмена и низких плотностях посадки (нагрузки) – 5-10 кг/м3 или 50-100 т/га. Нормативы ВНИИПРХ позволяют выращивать молодь форели до 16 кг/м3, а годовиков и двухлетков – 27-45 кг/м3. Высокое качество используемой воды может повысить уровень интенсификации форелеводства в 10-15 раз больше, чем наблюдается в настоящее время. При водообмене 10-15 раз в час реально получение 150-160 кг молоди и товарной форели с 1 м3 бассейнов. Это позволит выращивать 100 т товарной форели при нагрузке 10 кг/м3 на площади в 1 га, а при нагрузке 150 кг/м3 всего лишь на 0,07 га. Дальнейшее повышение уровня интенсификации форелеводства возможно при применении технического кислорода. Использование технического кислорода в форелеводстве – оксигенации открывает новые возможности интенсификации производства рыбы (рис.14) Таблица 14 Максимальная плотность посадки радужной форели при оксигенации, кг/м3
Качество воды имеет важное значение при выращивании форели всех возрастов. Вода не должна быть загрязнена химическими реагентами, быть прозрачной и умеренно жесткой, то есть содержать определенное количество солей магния и кальция. По ОСТ 15-282-83 вода для форелевых хозяйств должна соответствовать показателям таблицы 15, 16 и 17. Таблица 15 Показатели воды, поступающей в инкубационный форелевый цех
Таблица 16 Состав воды для форелевого хозяйства
Оценка загрязненности воды по химическим показателям
7.2. Отношение форели к некоторым абиотическим факторам средыАбиотические факторы среды оказывают существенное влияние на жизнедеятельность форели, и впервую очередь, температура воды. Температура воды один из универсальных и определяющих экологических факторов среды. Амплитуда, при которой живет форель различна для разных условий и составляет от 0,1 до 300С. Активность форели зависит от температуры воды как пойкилотермного организма. По отношению к температуре форель является стенотермной рыбой. Для форели оптимальная температура, как и для других рыб, зависит от возраста - икра 6 – 12,5, личинки, мальки 10 – 14 , сеголетки, годовики 14 – 16 , товарная рыба 14 – 180 С. Пороговая около 0,1, летальная 26 0С. При 18-200 и более – трудность поддержания газового режима, кислорода и активизация болезней. От температуры воды зависят сроки созревания, сроки нереста, продолжительность жизни. Растворенный в воде кислород. Его содержание тесно увязано с температурой воды. Оптимальные значения – 7 – 11 мг/л. В солоноватой и морской воде его растворяется меньше, чем в пресной. Чем моложе рыба, тем больше ей требуется растворенного кислорода. Для форели массой до 50 г необходимо 500 – 600 мг О2 кг. ч., а 100 – 200 г – 400-500 мг О 2 кг.ч. Содержание растворенного кислорода может колебаться в широких пределах в зависимости от температуры воды и других условий. Оптимальные условия дыхания у форели создаются при содержании кислорода на втоке 9-11, и не менее 5 мг О 2 /л – на вытоке. Форель реофильная и оксигенофильная рыба, живет при высоком парциальном давлении – хотя избыток, а также и недостаток кислорода могут вызвать у нее заболевание. Кислород оптимальный 9-11 мг л или 90-100% насыщения. Ночью, в предутренние часы наблюдается минимальное содержание кислорода. В солоноватой и морской воде содержание растворенного кислорода ниже, чем в пресной. Известно, что чем моложе форель, тем выше ее потребность в растворенном кислороде. Практически 1 л/с позволяет уверенно выращивать 60 кг товарной форели в год. Имеются различные способы улучшения содержания кислорода в воде. Скорость эмбрионального развития в сильной степени зависит от содержания растворенного кислорода. Форель может выдерживать 300-350% насыщения воды кислородом. Активная реакция среды рН (водородный показатель рН) - является показателем концентрации ионов водорода в воде и определяет: кислая, нейтральная или щелочная среда. Название происходит от английского power и химического знака ионов водорода Н. Определяется калориметрическим методом, в зависимости от окраски используют индикатор. Мерка – к 10 мл воды добавляют 4 капли индикатора. В зависимости от цвета – определяется рН - красный цвет – кислая среда, голубовато-фиолетовый – щелочная, зеленый цвет – нейтральная среда. Шкала от 4 до 10. Критическая для форели 9,2. Весной резко возрастает щелочность до 9 (гибель рыб). рН зависит от содержания Са в воде.. Нейтральное содержание рН –7. Благоприятные условия содержания рН – 6,5 – 8. Критические содержания ниже 6 и выше 8. Кислые воды болот содержат много гуминовых веществ (коричневая вода). Ручьевая форель выдерживает колебание рН от 4, до 9,5. Высокое содержание рН повышает воздействие ядовитого аммиака Nh4. При рН 5 форель теряет способность нормально размножаться. Углекислота, двуокись углерода, свободный диоксид (СО 2). Биопродуктивность водоема зависит от наличия двуокиси углерода . В большей концентрации углекислый газ ядовит для рыб. У форели уже при 30 мг СО2 / л наблюдается аритмия , угнетенное дыхание; при 50-80 мг/ л – нарушение равновесия, при 107 мг/л – плавание на боку. Углекислота (диоксид –СО2 ) или углекислый газ в природных водах содержится : 1) в свободном состоянии в виде газа, растворенного в воде – двуокись углерода;2) в виде ионов НСО3 – гидрокарбонат ионов3) в виде ионов СО3 карбонат – ионов. Аммиак, NН3. Рост карпа останавливается при содержании 0,06 мг NН3/л и задерживает рост молоди карпа. Имеется 2 формы аммиака - ионизированный аммиак или аммоний и неионизированный аммиак. Он оказывает токсическое действие, которое резко усиливается при повышении рН. Допустимая концентрация – 0,1 мг/ л. 3. Течение – носитель кислорода, удаляет продукты метаболизма (обмена ), остатки корма, экскременты. Равномерно распределяет корм. В лотках скорость течения 2-3 см/с. Крупная форель может преодолеть течение до 20 м.с. Известно, что большая скорость течения вызывает повышенный обмен и ухудшает рыбоводно-экономические показатели. Обычно течение не должно быть около 0,5 м/с. Содержание солей. Форель способна покрыть потребности в минеральных веществах из окружающей воды. Лучше если жесткая вода, чем мягкая. С возрастом концентрация солей может быть большей. Молодь хорошо растет при 3-6%0, неплохо переносит 9%0, 12-15%0 для двухлетней форели это уже нормальная соленость. При массе 100 г и более форель хорошо переносит соленость 30-35% 0. Пресные воды – содержат 1 г/л, солоноватые – 1 –15 г/л , соленые – 15-40 г/л минеральных солей. Источником поступления микроэлементов в рыбу является вода, растительность, естественный и искусственный корм. Жесткость – зависит от наличия солей Ca и Mg. За единицу жесткости принят градус жесткости: 1 немецкий градус – 10 Н= 10 мг Са в 1 л воды. 1 фрацузский градус – 10Ф = 10 мг СаСО3,. 1 английский градус – 10А = 10 мг СаСО3 в 700 г воды или 14,3 мг/л СаСО3. Жесткость бывает кальциевой и магниевой, а суммарная – общей жесткостью. Карбонатная жесткость СаСО3 – известь, характеризует концентрацию кальция и магния, а бикарбонатная – (СаСО3)2 – хорошо растворяется в воде до 900 мг/л или до 500. Жесткость, остающаяся в воде после кипячения, называется постоянной и выражается в мг-экв./л – 1 мг экв./л Са = 20,04 мг, 1 мг-экв./л Мg =12,16 мг. Карбонатная жесткость составляет 70-80% от общей жесткости. По степени жесткости воду делят на 6 классов (табл.18). Для форели важное значение имеет жесткость воды, которая должна в среднем составлять 8-12 Н0 (табл. 18) | ||||||
topuch.ru
Корма и кормление форели
Просмотров: 4633
В различных странах затраты, связанные с кормлением форели, при интенсивном товарном выращивании составляют 50—60 % всех производственных расходов, поэтому необходимо, чтобы кормление рыбы давало высокий экономический эффект.Энергетическая ценность корма и обмен веществ. Питательные вещества, содержащиеся в корме, должны не только поддерживать жизнедеятельность организма, но и способствовать его росту. С точки зрения энергетической ценности наиболее важными компонентами корма являются протеин, жир и углеводы.Наряду с белками, жирами и углеводами важное значение для поддержания жизненных функций имеют витамины и минеральные вещества. Если какое-либо необходимое вещество отсутствует или содержится в корме в недостаточном количестве, то это препятствует нормальному qbyHKILIMOHMpoBaHMKO opTaHM3Ma.Интенсивность обмена веществ зависит от различных факторов, в частности от размера рыбы — энергетическая потребность мелких рыб значительно выше, чем крупных. Так, при температуре 14 °С форель массой 1 г потребляет 480, а массой 100 г — 330 мг/(кг. ч) кислорода. Прием пищи повышает потребление энергии на 25%, а скорость течения в бассейне обусловливает интенсивность обмена веществ. При выращивании форели в стоячих водоемах наблюдается потеря тонуса мышц, что ухудшает качество товарной рыбы.Низкая концентрация растворенного кислорода приводит к увеличению скорости дыхания до определенного уровня, а затем происходит уменьшение скорости обмена и потребления пищи. При температуре 16 °C и выше содержание кислорода влияет на потребление пищи радужной форелью, поскольку максимальное количество пищи она потребляет при температуре 14 °C. Форель постепенно адаптируется к низкому содержанию кислорода, но при этом неэффективно использует корм. Мягкая вода может способствовать увеличению скорости обмена вследствие недостатка двухвалентных катионов. Высокое содержание аммиака и органических загрязнителей также увеличивает скорость обMeh3.У форели, испытывающей сильный стресс, интенсивность обмена может снижаться, что способствует сохранению энергии. Так как скорость обмена у радужной форели в значительной степени зависит от температуры, то энергетические потребности ее колеблются в зависимости от флюктуаций температуры воды.Радужная форель должна достичь товарной массы до наступления половой зрелости, поскольку в противном случае большое количество энергии будет израсходовано не на рост, а на формирование половых продуктов, в результате чего возрастает кормовой коэффициент.
Стартовые и продукционные корма. Обмен веществ у форели разного возраста имеет свои особенности, поэтому разработаны и выпускаются три группы кормов: стартовые (для молоди — с момента перехода на активное питание и до достижения ею массы 5 г), продукционные (для форели массой 5 г — до достижения ею товарной массы) и корма для производителей. В настоящее время в промышленном форелеводстве используют преимущественно сухие гранулированные корма (табл. 30).30. Состав стартовых и продукционных кормов для радужной форели, %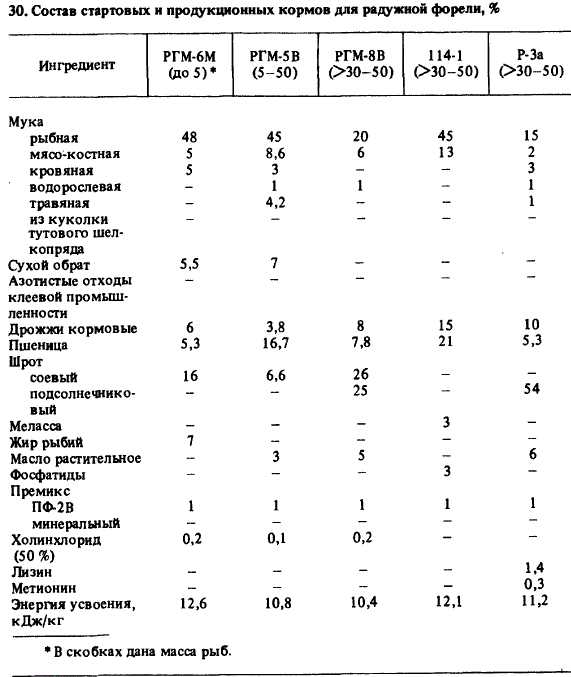
* В скобках дана масса рыб.Корма содержат более 40 различных структурных элементов, в том числе витаминные премиксы, минеральные вещества, антиокислители 122(витамин Е, цеолит клиноптилолит и др.), связывающие вещества (укрепляющие консистенцию корма), крилевый жир и пигмент антаксантин (для придания окраски мясу форели), лекарственные средства и ферментные препараты (для лечения и повышения перевариваемости питательных веществ корма). Кроме того, в корма вводят вещества, создающие привлекающий запах и положительное вкусовое ощущение, — аспарагиновую кислоту, тирозин, серин, глутаминовую кислоту, валин, MeTMOHСухие гранулированные корма выпускаются согласно ТУ 15-613—84 и ТУ 15-615—84 комбикормовыми заводами Минрыбхоза СССР в виде крупки и гранул (табл. 31).31. Соответствие между массой рыбы, качеством корма и частотой раздачи суточной нормыНомер частиц |Величина круп-|| Диаметр гра- Масса рыб, г Частота разда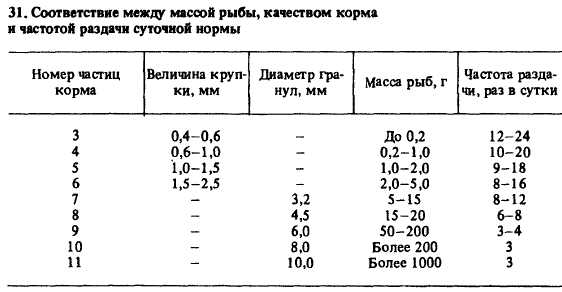
Необходимо отметить, что происходит постоянное подорожание традиционных компонентов кормов для форели, поэтому изыскание возможностей замены одних компонентов другими, особенно дефицитной рыбной муки, остается актуальной задачей.В связи с этим заслуживает внимания опыт ГДР по экономии гранулированных кормов при выращивании молоди форели за счет использования зоопланктона из естественных водоемов. Так, за 62 дня выращивания было использовано 5,5 т зоопланктона и 1 т сухого гранулированного корма, что позволило вырастить 200 тыс. шт. молоди радужной форели до достижения ею массы 1,5 г и получить значительную прибыль.Потребность радужной форели в питательных веществах при содержании в морской воде. Из-за необходимости регулирования ионного состава внутренней среды соленость следует рассматривать как фактор, постоянно требующий некоторых затрат энергии, связанных с активным транспортом ионов и необходимостью поддержания гомеостаза.Существует мнение, что причиной увеличения темпа роста радужной форели в соленой воде является лучшее усвоение корма, а не увеличение его потребления.123Установлено, что при одинаковой суточной дозе темп роста радужной форели при 16°С в пресной воде является максимальным и уменьшается по мере роста солености (до 20%o). В то же время при 8 °C темп роста радужной форели в морской воде соленостью 8 и 20%o выше, чем в пресной. Это говорит о том, что при содержании в морской воде соленостью 20 %о и более радужная форель особенно чувствительна к количеству и качеству корма. При содержании радужной фрpeли в морской воде соленостью 20 %о при 16 °С ей требуется больше пищи, чем при содержании в пресной или морской воде соленостью 8 %о при той же температуре.Выяснено, что радужная форель, акклимированная к определенной солености и температуре, потребует максимальное количество пищи в условиях промежуточной солености 15—28 %о, меньшее количество —в пресной воде и в воде соленостью 7,5 %o, а минимальное количество пищи — в воде соленостью 32,5 %o.Прекрасные результаты были получены при выращивании радужной форели в воде соленостью 10%o и 40%-ном содержании белка в сухом корме, а в воде соленостью 20 %o — при 45%-ном содержании белка.Испытание 57 кормовых смесей для радужной форели показало, что оптимальное содержание протеина в корме при выращивании в солоноватой воде составляет 46 %.В процессе выращивания радужной форели в морской воде наилучший прирост был получен при содержании протеина в корме от 47 μο 50 %.Для радужной форели, выращиваемой в морской воде, необходимо увеличить рацион до 150 % (по сравнению с рационом форели, выращиваемой в пресной воде).При экспериментальном выращивании радужной форели в бассейнахс использованием высокой плотности посадки (более 50—100 кг/м*) для соответствующих размерных групп и содержании кислорода на вытоке около 5 мг/л, как правило, не удавалось получить увеличения прироста радужной форели по сравнению с выращиванием в пресной воде в аналогичных условиях. Кроме того, считается, что плотность посадки рыбы может быть одной из причин, вызывающих повышенный отход при переводе в морскую воду.Прй выращивании в морской воде потребность радужной форели в аргинине понижается с 1,2 до 0,8 %, а скорость усвоения этой аминокислоты падает с 210 до 130 мг/(кг . сут). Для сеголетков радужной форели в морской воде требуется больше аргинина (1,2 % массы корма), чем в пресной (1 % массы корма). Соленость воды мало влияет на содержание в плазме крови незаменимых аминокислот, но содержание таурина, B-аланина, аспарагиновой и глутаминовой кислот и глицина у рыб, выращиваемых в соленой воде, значительно ниже, чем у рыб, выращиваемых в пресной воде.24Низкое содержание в корме жирных кислот вызывает повышенную смертность радужной форели в морской воде. Установлено, что в корм для радужной форели, выращиваемой в пресной и морской воде, следует добавлять рыбий жир.При морском выращивании радужная форель хорошо усваивает от 14 до 20% жира, содержащегося в корме. Темп роста радужной форели возрастает при увеличении содержания жира в корме до 20% даже при уменьшении содержания протеина в нем до 38%. Кроме того, было установлено, что рыба, получавшая корм с повышенным содержанием жира, обладает улучшенными вкусовыми качествами.Влажные и сухие корма. Для кормления лососевых рыб применяют как сухие (содержание влаги. 11—13%), так и влажные (содержание влаги около 50%) гранулированные корма.Влажные гранулированные корма имеют более мягкую консистенцию и приятный запах, что, видимо, привлекает рыбу и способствует быстрому потреблению корм
losos.arktikfish.com
Потребность рыбы в воде и кислороде
Просмотров: 70
Среди методов определения плотности посадки культивируемых рыб в условиях индуст-риального рыбоводства привлекает внимание метод, основанный на том, что концентра-ция рыбы или плотность посадки в единице рыбоводной емкости определяется количест-вом кислорода, необходимого для окисления суточной нормы корма. Как известно, спо-койная, не питающаяся рыба потребляет меньше кислорода, чем активная, питающаяся.
Потребление кислорода резко возрастает у питающейся рыбы за счет усиления обмена, окисления съеденного корма и выделения продуктов обмена. Возможное количество кор-ма, которое может быть использовано рыбой при конкретном количестве кислорода мо-жет быть вычислено следующим образом:
Х = (КН-КК)- 1,44- n / 220,
где: Х-количество корма, кг/сут.; Кн - начальное содержание кислорода в притекающей воде, мг/л; Кк - конечное минимальное содержание кислорода в вытекающей воде, 5 мг/л; n – количество воды, подаваемой в данную рыбоводную емкость, л/мин.; 1,44-количество воды в сутки при интенсивности подачи 1 л/мин., т; 220 - необходимое количество кисло-рода для усвоения рыбой 1 кг гранулированного корма с калорийностью 2600-2800 ккал/г (вычислено на основании эмпирических данных за 10 лет работы питомника Мак Ненни, США).
Установив количество корма, которое может быть использовано при данном количестве кислорода, определяется возможное количество рыбы в рыбоводной емкости и плотность посадки. При этом используют кормовые таблицы, например, таблицы ВНИИПРХ, в ко-торых показана суточная норма кормления форели в зависимости от массы тела и темпе-ратуры воды, то есть:
Возможное количество корма в сутки, кг/
количество количество корма в % к массе рыбы, кг
рыбы, кг
Например, температура воды, подаваемой в бассейны рыбоводного предприятия индуст-риального типа, равна 10 °С, масса рыбы 12 г, следовательно (по кормовым таблицам), для сухих гранулированных кормов суточная норма составит 2,6 % к массе рыбы, то есть:
Возможное 2,94кг/
количество 0,026 = 113,1 кг (9423 шт.)
рыбы, кг
Как видно, метод расчета плотности посадки рыбы основан на потребности в кислороде в зависимости от количества вносимого корма. Эта потребность в кислороде определена эмпирически и фактически учитывает зависимость потребления кислорода от температу-ры воды, размера рыбы и качества корма. Метод учитывает также и влияние продуктов обмена на способность рыбы использовать кислород в данных условиях кормления. Таким
образом, этот метод достаточно универсален. Однако он требует подробных данных о ве-личине суточного рациона в зависимости от температуры воды и массы рыбы. К настоя-щему времени эта зависимость изучена весьма тщательно, в основном для лососевых и карповых рыб. Она учитывает изменение физиологической активности при разной темпе-ратуре, следовательно, учитывает изменения общего обмена. Если при температуре 5 °С суточный рацион радужной форели массой 2-5 г составляет 2,2 %, то при температуре 10 °С - 3,3 %, а при 15 °С - 4,9 % от массы рыбы. Суточный рацион имеет обратную связь с массой тела рыбы. Если суточная норма для молоди лососей массой 2 г при температуре 10°С равна 4,2%, то для молоди массой 12-25 г - вдвое меньше. В связи с разнообразием условий на рыбоводных предприятиях, плотность посадки рыбы и количество воды на единицу выращиваемой рыбы рассчитывают не только на основании потребности рыбы в кислороде. В поступающей в рыбоводную емкость воде количество кислорода должно превышать потребность рыбы. Если при температуре воды 14-18 °С и близком к нормаль-ному насыщении (95 %) содержание кислорода составляет 8,93-9,75 мг/л (в среднем 9,34 мг/л), а на вытоке - 7 мг/л, то может быть использовано рыбой 2,34 мг кислорода из каж-дого литра притекающей в бассейны воды. Учитывая имеющиеся данные о расходе воды на 1 кг рыбы, количество поступающего с водой кислорода колеблется от 1193мг/кг-ч (при выращивании свободных эмбрионов) до 176мг/кг-ч в период товарного выращивания (табл. 92).
Таблица 92
Количество кислорода, поступающего в бассейны при эмпирически определенной
интенсивности подачи воды, и потребность молоди лососей в кислороде при
температуре 14-18 °С, насыщении 95% нормального и минимальное уровне 7 мг/л
Показатели Масса рыбы, г
0,140,251420160
Подача воды, л/мин, на 1 кг рыбы
Поступление кислорода, мг/кг • ч
Потребность в кислороде, мг/кг • ч
радужная форель
(Q = 0,601 W0,78)
лосось*
(Q = 0,712 W0,76)
Различия, % 8,5
1193
624
787
52 6,5
917
562
681
35 4
652
415
491
15 3
421
306
352
19 2,25
318
219
242
31 1,25
176
132
141
24
* При температуре 14-18 °С использован переводной коэффициент 1,45.
Вместе с тем потребность рыбы в кислороде, вычисленная по формулам Г.Г. Винберга и Л.П. Рыжкова меньше на 15-52%. Очевидно, этот избыток кислорода компенсирует по-вышение потребности его питающейся активной рыбой, а также покрывает затраты на окисление продуктов обмена. Не учитывается также кислород, поступающий из воздуха при активном перемешивании рыбой воды в бассейне. Эти расчеты показали, что в прак-тике рыбоводства потребность рыбы в кислороде значительно выше величин, определен-ных экспериментальным путем на примере спокойной, не питающейся рыбы.
Оптимальная плотность посадки и расход воды на единицу массы молоди лососевых рыб получены на основании выращивания при температуре воды от 14 °С до 18°С, то есть в условиях оптимума. Это дает основание с уверенностью использовать эти данные при вы-ращивании рыбы в условиях более низкой температуры воды, поскольку с понижением ее уменьшается интенсивность обмена. Соответственно этому уменьшается и потребность рыбы в кислороде. Следовательно, при более низкой температуре расход воды окажется избыточным. Поскольку расход воды на единицу продукции является экономическим фактором, представляется целесообразным уменьшать его величину в соответствии с уменьшением температуры воды. Это можно сделать, используя температурные коэффи-циенты для приведения значений обмена на любую температуру. Расчеты показали, что при снижении температуры от 14-18 °С до 3-5 °С потребность в воде снижается в 4-5 раз. Если при температуре 20 °С расчетный коэффициент равен 1, то при 14- 18°С-1,45, то есть потребность в воде снижается в 1,45 раза. Сначала необходимо определить расход воды при температуре 20 °С, затем, используя температурные коэффициенты, можно опреде-лить расход воды при других температурах.
Одновременно со снижением температуры воды, как известно, повышается растворимость в ней кислорода. Если при 20 °С нормальное насыщение воды кислородом составляет 9,02 мг/л, то при 1 °С - 14,25 мг/л. Следовательно, при снижении температуры повышается обеспеченность рыб кислородом и соответственно снижается потребность рыбы в воде. Чтобы учесть это снижение, введен кислородный коэффициент. Он показывает отношение
концентрации кислорода при интересующей нас температуре воды к концентрации кисло-рода при температуре 14-18 °С. При этой температуре количество растворенного в воде кислорода по средневзвешенному значению равно 9,82 мг/л (9,40-10,26 мг/л).
Принимая эту величину за единицу, при температуре воды выше 14-18 °С кислородный коэффициент будет менее единицы, при температуре воды ниже 14-18 °С- больше едини-цы. Разделив величины расхода воды на кислородный коэффициент, мы учтем снижение потребности в воде рыб, соответствующее повышению растворимости кислорода. Таким образом, если при температуре 14-18 °С, например, для свободных эмбрионов потреб-ность в воде составляет 8,1 л/мин., то при температуре 20 °С она повышается до 12,6 л/мин., а при температуре 3-5 °С - снижается до 1,3-1,7 л/мин. на 1 кг рыбы. Однако сле-дует учесть, что эмпирические данные о расходе воды при температуре 14-18 °С получе-ны в условиях насыщения воды кислородом до 95 %. Для удобства пользования расход воды приведен к насыщению 100 % (табл. 93).
Таблица 93
Потребность в воде молоди лососей в зависимости от температуры при
нормальном насыщении кислородом, л/мин, на 1 кг рыбы
Показатели Температура воды, °С
3456789101112
Температурный
коэффициент
Кислородный коэффициент
Стадия развития и масса
рыбы, г:
свободные эмбрионы 0,14(0,08-0,20)
личинки 0,25(0,15-0,35)
мальки до 1 г
мальки до 4 г
молодь посадочная,
покатная, смолты массой до 20 г
молодь посадочная,
товарная рыба массой
до 250 г 6,40
1,37
1,3
1,0
0,7
0,5
0,4
0,2
5,80
1,34
1,5
1,1
0,7
0,5
0,4
0,2
5,19
1,30
1,7
1,3
0,8
0,6
0,5
0,2
4,55
1,27
2,0
1,6
0,9
0,7
0,5
0,3 3,98
1,24
2,4
1,8
1,1
0,8
0,6
0,3
3,05
1,21
2,7
2,1
1,3
0,9
0,7
0,4
3,05
1,18
3,2
2,5
1,5
1,1
0,8
0,5
2,67
1,15
3,8
2,8
1,8
1,3
0,9
0,6
2,40
1,12
4,3
3,3
2,0
1,5
1,1
0,6
2,16
1,09
4,9
3,7
2,4
1,7
1,3
0,7
Продолжение таблицы 93
Показатели Температура воды, °С
131415161718192014-18
Температурный
коэффициент
Кислородный коэффициент
Стадия развития и масса
рыбы, г:
свободные эмбрионы
0,14(0,08-0,20)
личинки 0,25(0,15-0,35)
мальки до 1 г
мальки до 4 г
молодь посадочная,
покатная, смолты массой до 20 г
молодь посадочная,
товарная рыба массой
до 250 г 1,94
1,07
5,6
4,3
2,7
2,0
1,5
0,8
1,74
1,04
6,5
4,9
3,0
2,3
1,7
1,0
1,57
1,02
7,2
5,6
3,4
2,5
1,9
1,0 1,43
1,00
8,2
6,3
3,8
2,8
2,2
1,2
1,31
0,98
9,1
6,9
4,3
3,2
2,5
1,3
1,20
0,96
10,1
7,7
4,7
3,5
2,7
1,5
1,09
0,94
11,5
8,7
5,3
3,9
3,0
1,6
1,00
0,92
12,6
9,6
6,0
4,5
3,4
1,9
1,45
1,00
8,1
6,2
3,8
2,8
2,1
1,2
Следовательно, если в конкретном рыбоводном предприятии индустриального типа в ры-боводные бассейны поступает вода с концентрацией кислорода менее 100% насыщения, табличные данные увеличиваются следующим образом:
V= 100 n/М,
где: V - искомый расход воды, л/мин, на 1 кг рыбы; n - расход воды при 100%-ном насы-щении воды кислородом; М-насыщение воды кислородом в конкретном бассейне (во всем предприятии), % от нормального.
Потребность в воде при разной температуре, представленная графически, выглядит в виде параболических кривых, которые при понижении температуры воды имеют тенденцию к выпрямлению.
В практических целях представляет также интерес не расход воды на 1 кг массы выращи-ваемой рыбы, но, наоборот, возможная посадка рыбы (в кг) на 1 л/мин, подаваемой воды. Как видно, это взаимообратные величины. Для свободных эмбрионов, например, при тем-пературе 14-18 °С требуется расход воды 8,2 л/мин, на 1 кг, в то время как в расчете на 1 л/мин, подаваемой воды можно посадить всего лишь 0,12кг свободных эмбрионов. В про-цессе выращивания молоди эти величины сближаются.
Таким образом, для определения плотности посадки рыбы и интенсивности водообмена в бассейнах рыбоводного предприятия индустриального типа следует использовать эмпи-рические методы. Определение оптимальной плотности посадки рыб различных возрас-тных групп позволяет вычислить необходимый водообмен.
biblio.arktikfish.com
КИСЛОРОД И ЕГО ПОТРЕБЛЕНИЕ РЫБОЙ
Просмотров: 5421
Для всех культивируемых водных объектов необходим кислород, который им приходитсяизвлекать из воды. Основным источником пополнения кислорода в природной воде являетсяатмосфера и фотосинтезирующие растения. В атмосфере содержится 21% кислорода, а содер-жание кислорода в воде зависит от различных факторов, в частности, от температуры воды исодержания в ней ионов хлора. Для каждой температуры и солености (хлорности) может бытьустановлено равновесное 100% насыщение атмосферным кислородом. В холодной пресной во-де насыщение выше, чем в более теплой и насыщенной ионами хлора. Равновесное насыщениевесьма важный показатель, по которому оценивается несущая способность воды по кислороду.Для практического пользования эта информация представлена в виде табл.12.Таблица 12.
Если по каким-либо причинам насыщение воды в водоеме оказывается ниже равновесногонасыщения, то кислород воздуха переносится в воду из-за возникающего градиента концентра-ций.
Скорость процесса переноса кислорода тем выше, чем выше градиент концентрации, тоесть чем выше разница между равновесным и фактическим насыщением воды. При приближе-нии фактического насыщения воды к равновесному скорость процесса падает. При пресыщениеводы кислородом выше равновесного насыщения идет процесс потери кислорода из воды(диффузия). Процессы насыщения и диффузия идут через поверхность раздела сред во-да/воздух, поэтому, чем больше поверхность раздела, тем интенсивнее идет процесс. Поверх-ность раздела может быть искусственно увеличена за счет разбрызгивания или барботированиявоздухом. Создание турбулентного течения воды также увеличивает скорость процесса, так какувеличивается градиент концентрации на границе раздела при перемешивании слоев воды оразличной степенью насыщения. Процессы диффузии кислорода в неподвижной воде настолькомало влияют на уровень насыщения различных слоев воды, что ими можно пренебречь.Фотосинтез - второй основной источник кислорода для водных систем. Кислород являетсяпобочным продуктом фотосинтеза растений. Однако, фотосинтез и продуцирование кислородапроисходят только при наличии света. Те водоросли, которые в дневное время выделяют ки-слород, в ночное время его поглощают. В солнечные дни количество кислорода, выделяемоговодорослями в воду, может быть настолько большим, что уровень насыщения воды значитель-но превышает уровень равновесного насыщения. В этом случае кислород выделяется из воды ватмосферу. Известны случаи пресыщения воды до 300% от равновесного значения. Макси-мальная концентрация кислорода бывает в середине дня или несколько позднее, а минимальнаяконцентрация - на рассвете.Потребление водорослями кислорода в темное время суток может снизить его концентрациюв воде до критического для культивируемых объектов значения. Особую опасность представ-ляют тихие безветренные ночи, когда пополнение воды кислородом воздуха затруднено. Кон-троль концентрации кислорода в открытых водоемах следует проводить в утренние часы.НОРМА КОНЦЕНТРАЦИИ КИСЛОРОДА В ВОДЕ РЫБОВОДНЫХ УСТАНОВОК.В естественных открытых водоемах, в результате процесса фотосинтеза водорослей, верхняяграница насыщения может оказаться равной 300% от равновесного насыщения. При этом неотмечено гибели рыбы или каких-либо других отрицательных последствий.Снижение концентрации кислорода до 60% от равновесного насыщения практически не ска-зывается на росте и использовании кормов форелью и карпом. Временное снижение концентра-ции кислорода ниже 60% приводит к стрессам на несколько дней, рыба прекращает брать корм.При дальнейшем снижении концентрации кислорода наступает гибель рыбы. Нижние порого-вые концентрации кислорода зависят от вида и массы тела рыбы, а также от температуры воды.Пороговые концентрации кислорода являются важной информацией при выполнении операцийпо перевозке рыбы. В этот момент рыба не питается, а единственной задачей рыбовода являетсясохранение рыбы в живом виде. Пороговые концентрации кислорода в воде для различных ви-дов рыб приведены в табл.13.Оптимальные концентрации кислорода для рыбы подбираются в зависимости от стадии ееразвития; икра, личинки, молодь, товарная рыба. На стадии инкубации икры требования куровню концентрации кислорода жестче, чем на других стадиях. Например, при инкубации ик-ры карпа снижение концентрации кислорода до 6,5 мг/л, что составляет 70% равновесного на-сыщения, оказывается летальным. В период усиленного развития икринки, во время закладкиосновных органов и тканей зародыша, даже 100% насыщение воды 8,5 мг/л становится крити-ческим, часть икры погибает. Повышение концентрации кислорода до 25 - 35 мг/л не сказыва-ется на скорости развития икры. Однако, при столь высокой концентрации кислорода уменьша-ется отход икры. В дальнейшем развитии молодь, полученная из икринок, инкубированных приповышенной концентрации кислорода, развивается и набирает массу быстрее молоди, по лу-ченной из икры, инкубированной при 100% насыщении воды.45Таблица 13
На стадии личинки, когда организм рыбы претерпевает морфологические преобразования,требования к уровню концентрации кислорода не менее жесткие. Концентрация кислородадолжна поддерживаться на уровне 100% насыщения, хотя более высокие концентрации не при-носят очевидного вреда, если они не связаны с газопузырьковой опасностью.На более поздних стадиях развития молоди и выращивания товарной рыбы оптимальныеконцентрации кислорода лежат в пределах от 90 до 200% насыщения. Дальнейшее увеличениеконцентрации эффекта не дает. Очевидно, что при высоких концентрациях кислорода усвоениекорма рыбой идет эффективнее, а затраты энергии на дыхание и поиск зон с повышенной кон-центрацией кислорода сводятся к минимуму. О недостатке кислорода можно судить по поведе-нию рыбы. На недостаток кислорода указывает концентрация рыбы на вводе воды в бассейн,выпрыгивание, скопление рыбы в верхних слоях воды.ПОТРЕБЛЕНИЕ КИСЛОРОДА РЫБОЙ.Потребление кислорода рыбой зависит от стадии развития, массы тела, температуры и коли-чества потребленного корма. По мере развития эмбриона потребность в кислороде возрастает.При инкубировании икры в условиях цеха потребность в кислороде удовлетворяется за счет во-ды, протекающей через инкубатор. При перевозке икры на стадии глазка потребность в кисло-роде покрывается тем запасем кислорода, который помещен в контейнер для перевозки икры. Сростом температуры потребление кислорода эмбрионами растет. Рост потребления кислородаэмбрионами осетровых приведен в табл.14.Таблица 14
При переходе личинок на питание внешним кормом по мере развития и роста рыбы удельноепотребление кислорода на единицу массы рыбы падает. Одновременно сохраняется зависи-мость уровня удельного потребления кислорода в функции температур в пределах температур,возможных для разведения. Эта зависимость иллюстрируется потреблением кислорода лосося-ми и форелью в диапазоне температур от 7,2 до 21,1 оС для рыб массой до 1000 г. (рис.21).Рис. 21. Потребление кислорода в зависимости от массы рыбы и температуры.Потребление кислорода в течение суток неравномерно. Максимальное потребление ки-слорода у форели наблюдается чрез 10 - 15 мин после внесения корма и снижается через 1 - 1,5часа. Уровень потребления кислорода колеблется в пределах 1,2 - 2 раза. Аналогичная картинанаблюдается при кормление карпа. По этой причине все данные о потреблении кислорода при-водятся в расчете на сутки. Для практических расчетов потребности кислорода для рыбы расчетведется по потребленному ей корму. В пределах температур оптимальных для форели потреб-ление кислорода пропорционально потреблению корма. На 1 кг сухого гранулированного кормарыбой расходуется 0,22 кг кислорода. Такой расчет применим только для условий нормальноготехнологического процесса выращивания рыбы. Действительно, при прекращение кормлениярыбы потребление кислорода не станет равным нулю.
biblio.arktikfish.com
КИСЛОРОД И ЕГО ПОТРЕБЛЕНИЕ РЫБОЙ
Просмотров: 5422
Для всех культивируемых водных объектов необходим кислород, который им приходитсяизвлекать из воды. Основным источником пополнения кислорода в природной воде являетсяатмосфера и фотосинтезирующие растения. В атмосфере содержится 21% кислорода, а содер-жание кислорода в воде зависит от различных факторов, в частности, от температуры воды исодержания в ней ионов хлора. Для каждой температуры и солености (хлорности) может бытьустановлено равновесное 100% насыщение атмосферным кислородом. В холодной пресной во-де насыщение выше, чем в более теплой и насыщенной ионами хлора. Равновесное насыщениевесьма важный показатель, по которому оценивается несущая способность воды по кислороду.Для практического пользования эта информация представлена в виде табл.12.Таблица 12.
Если по каким-либо причинам насыщение воды в водоеме оказывается ниже равновесногонасыщения, то кислород воздуха переносится в воду из-за возникающего градиента концентра-ций.
Скорость процесса переноса кислорода тем выше, чем выше градиент концентрации, тоесть чем выше разница между равновесным и фактическим насыщением воды. При приближе-нии фактического насыщения воды к равновесному скорость процесса падает. При пресыщениеводы кислородом выше равновесного насыщения идет процесс потери кислорода из воды(диффузия). Процессы насыщения и диффузия идут через поверхность раздела сред во-да/воздух, поэтому, чем больше поверхность раздела, тем интенсивнее идет процесс. Поверх-ность раздела может быть искусственно увеличена за счет разбрызгивания или барботированиявоздухом. Создание турбулентного течения воды также увеличивает скорость процесса, так какувеличивается градиент концентрации на границе раздела при перемешивании слоев воды оразличной степенью насыщения. Процессы диффузии кислорода в неподвижной воде настолькомало влияют на уровень насыщения различных слоев воды, что ими можно пренебречь.Фотосинтез - второй основной источник кислорода для водных систем. Кислород являетсяпобочным продуктом фотосинтеза растений. Однако, фотосинтез и продуцирование кислородапроисходят только при наличии света. Те водоросли, которые в дневное время выделяют ки-слород, в ночное время его поглощают. В солнечные дни количество кислорода, выделяемоговодорослями в воду, может быть настолько большим, что уровень насыщения воды значитель-но превышает уровень равновесного насыщения. В этом случае кислород выделяется из воды ватмосферу. Известны случаи пресыщения воды до 300% от равновесного значения. Макси-мальная концентрация кислорода бывает в середине дня или несколько позднее, а минимальнаяконцентрация - на рассвете.Потребление водорослями кислорода в темное время суток может снизить его концентрациюв воде до критического для культивируемых объектов значения. Особую опасность представ-ляют тихие безветренные ночи, когда пополнение воды кислородом воздуха затруднено. Кон-троль концентрации кислорода в открытых водоемах следует проводить в утренние часы.НОРМА КОНЦЕНТРАЦИИ КИСЛОРОДА В ВОДЕ РЫБОВОДНЫХ УСТАНОВОК.В естественных открытых водоемах, в результате процесса фотосинтеза водорослей, верхняяграница насыщения может оказаться равной 300% от равновесного насыщения. При этом неотмечено гибели рыбы или каких-либо других отрицательных последствий.Снижение концентрации кислорода до 60% от равновесного насыщения практически не ска-зывается на росте и использовании кормов форелью и карпом. Временное снижение концентра-ции кислорода ниже 60% приводит к стрессам на несколько дней, рыба прекращает брать корм.При дальнейшем снижении концентрации кислорода наступает гибель рыбы. Нижние порого-вые концентрации кислорода зависят от вида и массы тела рыбы, а также от температуры воды.Пороговые концентрации кислорода являются важной информацией при выполнении операцийпо перевозке рыбы. В этот момент рыба не питается, а единственной задачей рыбовода являетсясохранение рыбы в живом виде. Пороговые концентрации кислорода в воде для различных ви-дов рыб приведены в табл.13.Оптимальные концентрации кислорода для рыбы подбираются в зависимости от стадии ееразвития; икра, личинки, молодь, товарная рыба. На стадии инкубации икры требования куровню концентрации кислорода жестче, чем на других стадиях. Например, при инкубации ик-ры карпа снижение концентрации кислорода до 6,5 мг/л, что составляет 70% равновесного на-сыщения, оказывается летальным. В период усиленного развития икринки, во время закладкиосновных органов и тканей зародыша, даже 100% насыщение воды 8,5 мг/л становится крити-ческим, часть икры погибает. Повышение концентрации кислорода до 25 - 35 мг/л не сказыва-ется на скорости развития икры. Однако, при столь высокой концентрации кислорода уменьша-ется отход икры. В дальнейшем развитии молодь, полученная из икринок, инкубированных приповышенной концентрации кислорода, развивается и набирает массу быстрее молоди, по лу-ченной из икры, инкубированной при 100% насыщении воды.45Таблица 13
На стадии личинки, когда организм рыбы претерпевает морфологические преобразования,требования к уровню концентрации кислорода не менее жесткие. Концентрация кислородадолжна поддерживаться на уровне 100% насыщения, хотя более высокие концентрации не при-носят очевидного вреда, если они не связаны с газопузырьковой опасностью.На более поздних стадиях развития молоди и выращивания товарной рыбы оптимальныеконцентрации кислорода лежат в пределах от 90 до 200% насыщения. Дальнейшее увеличениеконцентрации эффекта не дает. Очевидно, что при высоких концентрациях кислорода усвоениекорма рыбой идет эффективнее, а затраты энергии на дыхание и поиск зон с повышенной кон-центрацией кислорода сводятся к минимуму. О недостатке кислорода можно судить по поведе-нию рыбы. На недостаток кислорода указывает концентрация рыбы на вводе воды в бассейн,выпрыгивание, скопление рыбы в верхних слоях воды.ПОТРЕБЛЕНИЕ КИСЛОРОДА РЫБОЙ.Потребление кислорода рыбой зависит от стадии развития, массы тела, температуры и коли-чества потребленного корма. По мере развития эмбриона потребность в кислороде возрастает.При инкубировании икры в условиях цеха потребность в кислороде удовлетворяется за счет во-ды, протекающей через инкубатор. При перевозке икры на стадии глазка потребность в кисло-роде покрывается тем запасем кислорода, который помещен в контейнер для перевозки икры. Сростом температуры потребление кислорода эмбрионами растет. Рост потребления кислородаэмбрионами осетровых приведен в табл.14.Таблица 14
При переходе личинок на питание внешним кормом по мере развития и роста рыбы удельноепотребление кислорода на единицу массы рыбы падает. Одновременно сохраняется зависи-мость уровня удельного потребления кислорода в функции температур в пределах температур,возможных для разведения. Эта зависимость иллюстрируется потреблением кислорода лосося-ми и форелью в диапазоне температур от 7,2 до 21,1 оС для рыб массой до 1000 г. (рис.21).Рис. 21. Потребление кислорода в зависимости от массы рыбы и температуры.Потребление кислорода в течение суток неравномерно. Максимальное потребление ки-слорода у форели наблюдается чрез 10 - 15 мин после внесения корма и снижается через 1 - 1,5часа. Уровень потребления кислорода колеблется в пределах 1,2 - 2 раза. Аналогичная картинанаблюдается при кормление карпа. По этой причине все данные о потреблении кислорода при-водятся в расчете на сутки. Для практических расчетов потребности кислорода для рыбы расчетведется по потребленному ей корму. В пределах температур оптимальных для форели потреб-ление кислорода пропорционально потреблению корма. На 1 кг сухого гранулированного кормарыбой расходуется 0,22 кг кислорода. Такой расчет применим только для условий нормальноготехнологического процесса выращивания рыбы. Действительно, при прекращение кормлениярыбы потребление кислорода не станет равным нулю.
biblio.arktikfish.com

















